Экспрессия генов в ЦНС беспозвоночныхСтраница 2

Наконец, классический объект генетических исследований - дрозофила - позволил получить обширную информацию о сложных процессах онтогенеза центральной нервной системы. Эту информацию можно обобщенно представить в виде модели многоступенчатой генетической детерминации нейрогенеза, основные положения которой, по-видимому, сохраняют силу и для позвоночных животных. В соответствии с этой моделью существует четыре класса генов, регулирующих последовательные этапы нейрогенеза. Первый, наименее изученный класс, включает гены, ответственные за преобразование части клеток недифференцированной вентральной эктодермы в предшественники нервных клеток. Ко второму классу относятся гены, детерминирующие свойства клеток-предшественников в соответствии с их положением в той или иной части ЦНС. Среди этих генов - многие из так называемых гомейотических или сегментарных генов, которые детерминируют также и формирование общего плана строений организма в целом. При этом одни и те же гены обычно детерминируют и формирование определенного сегмента организма, и специфические свойства локализованного в нем сегмента ЦНС. Заметим, однако, что сегментарная и нейроспецифическая экспрессия гомейотических генов контролируется разными механизмами. Гены третьего класса детерминируют свойства индивидуальных нейробластов в зависимости от их локализации внутри данного сегмента ЦНС. Представителем этого класса, по-видимому, является ген prospero, экспрессирующийся только в определенных нейробластах каждого сегмента ЦНС, но не в образующихся из них нейронах. Мутации этого гена приводят к выпадению из развивающейся нервной системы нескольких специфических клеточных линий. Наконец, гены четвертого класса детерминируют индивидуальные свойства каждого из нейронов, образующегося при делении одного нейробласта. Представителями этого класса являются гены fushi tarazu и even-skipped. Интересно, что один и тот же ген может участвовать в контроле разных этапов нейрогенеза. Так, ген ftz является типичным гомейотическим геном, участвующим в формировании общего сегментарного плана строения ЦНС и организма в целом, т.е. входит во второй класс генов. На более поздних этапах он участвует в детерминации свойств индивидуальных нейронов: в каждом сегменте ЦНС он экспрессируется в группе из 30 идентифицируемых нейронов, являющихся потомками - 8 нейробластов. При этом ни в одном из нейробластов ген ftz не экспрессируется, а процент экспрессирующих его клеток в потомстве индивидуальных нейробластов варьирует от 0 до 100%. Заметим, однако, что сегментарная и нейроспецифическая экспрессия гена ftz контролируется разными механизмами. Еще один фундаментальный вывод описанных исследований состоит в том, что индивидуальная специфичность элементов нервной системы определяется не уникальными генами, количество которых заведомо меньше числа самих элементов, а уникальными комбинациями взаимодействующих генов, надежность которых обеспечивается чертами функциональной избыточности.
Гены, ответственные за формирование специфического фенотипа функционально зрелых нейронов, изучены главным образом для относительно просто устроенных нейронных структур, например сетчатки глаза дрозофилы. Каждый элемент сетчатки - омматидий - состоит из 8 четко различимых по морфологии и локализации фоточувствительных нейронов и 12 ненейрональных клеток. Дифференцировка нейронов в развивающихся омматидиях протекает в строго фиксированной последовательности. Недифференцированные клетки в зачатках омматидиев являются эквипотенциальными, а судьба дифференцирующихся клеток определяется не их происхождением, а индуцирующими стимулами со стороны соседствующих с ними ранее дифференцированных клеток.
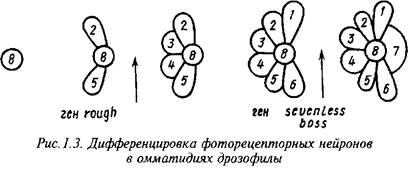
Некоторые из генов, ответственных за генерацию и рецепцию этих стимулов, хорошо изучены: ген seveniess кодирует мембранный рецептор, необходимый для индукции дифференцировки нейрона R7, а ген bride of sevenless - за образование в нейроне R8 лиганда, действующего на этот рецептор. Ген seven in absentia кодирует ядерный белок, интерпретирующий сигнал, воспринимаемый рецептором sevenless. Ген rough кодирует транскрипционный фактор, необходимый для возникновения в нейронах R2 и R6 сигнала, индуцирующего дифференцировку соседних клеток в нейроны R3 и R4. Другой транскрипционный фактор, необходимый для дифференцировки нейронов Rl, R3, R4 и R6, кодируется геном seven-up. Ключевым элементом в реализации генетических программ функционального созревания фоторецепторных нейронов является ДНК-связывающий белок, кодируемый геном glass, а специфическими для индивидуалъньгх фоторецепторных нейронов компонентами этих программ - гены, кодирующие разные формы фоторецепторных белков.
Популярные статьи:
Механизм взаимодействия излучений человека и окружающей среды и возможности
медицинской диагностики и лечения
Из тех, что мир прошли в вдоль и поперек,
Из тех, кого Творец на поиски обрек,
Нашел ли хоть один хоть что-нибудь такое,
Чего не знали мы и что пошло нам впрок?
Омар Хайям
Ощущения - это обман наших чувств.
Р. Декарт
Внешние электр ...
Формы акации белой
Основной задачей лесного семеноводства является получение семян с высокими наследственными свойствами, поэтому семеноводство акации должно строиться на селекционной основе, т. е. речь должна идти не о семеноводстве вообще, а о семеноводст ...
Выделение и очистка белковых препаратов. Очистка
С-белка, Х-белка и Н-белка
Х-белок, С-белок и Н-белок очищали по методу (Offer et al., 1973). Фракции Х-, С- и Н-белков, полученные при хроматографической очистке миозина скелетных мышц на колонке с носителем DEAE-Sephadex А-50, концентрировали сульфатом аммония до ...